Give an account of the following:
Q. Give an account of the following:
(i) Antheridiophore of Marchantia
When the thallus of Marchantia attains maturity it bears upright stalked umbrella like structures at the apices of certain lobes. These structures are the antheridiophores which bear antheridia and the archegoniophores which bear archegonia.
The male branch or antheridiophore is a modified branch of the thallus. It has a short stalk with an eight-lobed flat peltate disc at its apex. The eightlobed condition of the disc is due to the repeated forking of the young branch. As a result of these divisions very often eight lobes are formed though occasionally their number may be reduced to four in certain species. The stalk has two grooves on the side where rhizoids and scales are given out and this side corresponds to the ventral side of the thallus.
The peltate disc is slightly convex. In a vertical section it exhibits the same structure as that of the thallus, of which it is a modified branch. On the upper side, there is a single-layered epidermis which has many barrel-shaped pores. Each pore opens into a photosynthetic chamber. Alternating with these pores are other openings which lead into big antheridial cavities.
Each cavity has a single short-stalked antheridium. On ripening, each antheridium has many androcytes.
(ii) Equisetum Strobilus
Each cone or strobilus is a terminal structure consisting of a thick somewhat conical axis from which a whorl of about twenty densely arranged peltate sporangiophores is given off at right angles. Because of mutual pressure, the mature sporangiophores are typically hexagonal in surface view. These fit closely together and form a protective covering for the sporangia which are on the underside of this plate. The lower most whorl is sterile and forms the rudimentary sheath in the forms of a collar. At the base of the cone there is a ring like toothed outgrowth, the annulus.
The nature of these sporangiophore has led to divergent views. Some regard it as a lateral branch developing a whorl of sporangia, others prefer regarding it as the fertile ventral lobe of a dorsoventrally divided sporophyll, a few consider it as the stalk of a raised and divided sporangium, some call it a sporophyll, whereas others regard it as an organ sui-generis.
Mild, Bower, Goeble, Kashyap, Tschudy and Sinha mentioned some abnormalities in the cones of Equisetum. Sinha collected some abnormal cones of E. debile from Meerut. One of the specimen was found to have the cone divided into two segments by a narrow transverse constriction having a fertile sheath comparable to the annulus while another specimen showed a short proliferated shoot above the cone.
Two views have been advanced to account for the morphological nature of the strobilus. The most widely accepted view is that of Brown who regards strobilus to be formed of a short axis consisting of nodes and internodes. The position of the sporangiophores marks the node. This view is opposed by Barrat.
(iii) Spike of Ophioglossum
A pyramid-like apical cell with four cutting faces gives rise to a conical structure of meristematic tissue that represents a very young fertile spike. This conical outgrowth turns upwards. According to Bower, the four quadrants cut just back of the apex of the spike corresponding to the four cutting faces of the apical cell. These quadrants are oriented in such a way that two lie in a plane perpendicular to the sterile lamina, and two in a plane parallel to it.
The fertile spikes arise from the basal portion of the sterile leaf. The fertile spike may easily be differentiated from the sterile segment. It is usually erect and stalked.
The sporangia are found within the tissue of the fertile spike. The sporangia are found to be arranged on the margins of the spike in two longitudinal rows. The number of sporangia varies from 6-20 on a fertile spike.
The morphological nature of the fertile spike is controversial. According to Bower it is a single septate sporangium but this hypothesis has been rejected later on by Bower himself. The pteridologists of modern days are in agreement that it is pinna like in nature. It has been held by Goebel that the spike represents a single pinna. But according to Chrysler it is more probable that the fertile spike represents the fusion of the two basal pinnae of a leaf. This interpretation has been supported on the basis of vascular supply. The fertile spike has the same vascular supply as would go to a pair of pinnae.
(iv) Sporophyte of Polytrichum
The diploid (2x) zygote or oospore is the first structure of the sporophyte. The zygote is located in the nter of archegonium situated at the top of the female plant. The oospore increases in size and develops into a mature sporogonium. So the growth of female plant stops and new sporophytic structure sporogonium comes into existence. The mature sporogonium consists of a foot, a long seta and a capsule. The foot is embedded in the tissue of the apex of an archegonial branch and consists of a thin walled parenchymatous cells. Just above the foot and continuous with it is a long slender seta which bears a capsule at its apex. Actually it is the stalk of the capsule.
At the base of the capsule seta enlarges to form the apophysis which is marked off from the sporogenous portion by a groove. The apophysis has a distinct epidermis with stomata which are confined to the region of groove only. Inside the epidermis there is a loosely arranged mass of chlorenchymatous tissue and thus it is the main photosynthetic region. In the ntre it is the conducting strand continuous with the columella above.
Above the apophysis is the capsule. The capsule is erect in position and completely covered by the brown calyptra with a hairy surface even when nearly mature.
The body of capsule is differentiated into the tower spore bearing portion called theca and a sterile conical portion called operculum.
Theca is covered by capsule wall of several layers. The outermost layer of the wall consists of a compactly arranged cells with their outer walls thickened. It is known as epidermis. The apex of the capsule consists of an operculum which is a conical lid with a long beak.
(v) Salient features of Archegoniophore of Marchantia
The archegoniophore consists of a stalk with a eight lobed apical disc.
The stalk of archegoniophore resembles antheridiophore in morphology and anatomy. It is slightly longer than antheridiophore. It may be 2-3 inches long.
The disc of archegoniophore becomes lobed in the following manner. When the archegoniophore is still very young, its apex forks and becomes swollen and dichotomy repeated in quick succession as a result of which the young archegoniophore ultimately becomes a rosette like eight lobed disc. The apical cells of each lobe cut off segments on dorsal and ventral faces. The archegonia are formed in acropetal succession from the segments cut off on the dorsal face. Soon eight groups of archegonia develop on the upper surface of the disc, corresponding to eight growing points of disc.

Fig. Marchantia-V. S. of archegoniophore bearing hanging archegonia.
After fertilization the stalk of archegoniophore begins to elongate and side by side there is a conspicuous overgrowth in the sterile central part of the disc with the result the marginal apical region of the disc alongwith the group of archegonia are pushed over the lower surface.
Finally the growing apices are turned downward and comes close to the stalk of archegoniophore. After curvature, one layered plate of tissue develop on either side of each group of archegonia. Thus each group of archegonia (12-15) is enclosed by two lipped pendent fringed sheaths known as perichaetium which hangs down vertically from the underside of the lobe of disc.
(vi) Prothallus of Psilotum
The apical cell forms a mass of cells through repeated divisions in which endophytic fungus penetrates in early stage. This mass of cells forms the young prothallus which increases in size rapidly till it attains the full size. The prothallus is a gametophyte.
The mature prothallus is more or less cylindrical, elongated subterranean body which is radially symmetrical and imperfectly dichotomously branched. The size of prothallus is upto 20 mm long and 2 mm in diameter. It is found growing in the crevices of rocks or on tree trunks in natural condition. It is colourless or yellowish brown.

Fig. Entire prothallus showing all structure
It is densely covered with numerous dark brown stiff hair like rhizoids. It is saprophytic and depends upon the cooperation of a mycorrhizal endophytic fungus for its nutrition which gains its entrance through rhizoids and invades nearly all cells of prothallus. The prothallus is mostly composed of colourless hexagonal cells. The cells are highly cutinized along their outer and radial wall. The rhizoids are developed from the outer layer at frequent intervals. There is an interrupted vascular supply in prothallus. The stele is composed of scalariform or scalariform reticulate tracheids surrounded by phloem. This interrupted vascular strand is essentially protostele and it is interrupted by intervening parenchyma. The presence of such well differentiated vascular strand in prothallus is an usual character. Cytologically the prothallus is diploid.
(vii) Elaters of Bryophytes
Elaters found in capsules of bryophytes play an important role in their bursting. The hygroscopic nature of elaters allow them to absorb moisture as a result of which they swell. The swelling of elaters exerts pressure on the walls of capsules, which ultimately leads to bursting of the capsules.
Elaters are tubular cells with spiral thickenings and often help in spore release. Elaters do not work in the same way in all species and there are some species where elaters play little or no role in spore release.
An elater is a cell (or structure attached to a cell) that is hygroscopic, and therefore will change shape in response to changes in moisture in the environment. Elaters come in a variety of forms, but are always associated. with plant spores. In many plants that do not have seeds, they function in dispersing the spores to a new location.
The sporophyte of a Bryophyte, generally consisting of a foot, seta and capsule. The mature capsule is filled with spores mingled with sterile cells or elaters and opens by splitting into valves.
(viii) Gemma cup of Marchantia
The gemmae which are the special reproduction bodies, produced in large numbers in small gemmae cups. Each gemma has a small stalk and a multicellular discoid body. It is thick in the middle and thin at margins. There are two notches on the lateral sides which are diagrammatically opposite, while few possess oil. There are some rhizoidal cells also.
Each gemmae after falling on the soil and on the approach of favourable conditions develop into two thallus of Marchantia gametophyte. First of all two apical cells in the two notches become active and begin to grow and develop in the same manner as those at the growing tip of a branch. Due to the activities of these two cells in opposite direction, two young thalli are formed simultaneously. The rhizoids are formed from the rhizoidal cells. Ultimately the gemma exhausted and dies and the two young, thalli get separated. These thalli developed into two gametophytes of Marchantia.

Fig. V. S. of Marchantia thallus passing through a gemma cup
Development of gemmae takes place in the gemma cups on the dorsal surface of thallus. Gemma cups are formed by rapid growth of neighbouring cells thus assuming a cup like structure. Any cell of the floor of gemma cup functions as gemma initial which divides by a transverse division into a stalk cell and gemma cell. The stalk cell directly forms a stalk while the gemma cell by repeated transverse and vertical division forms a gemma. Afterwards by periclinal division in the central cells it becomes several cells in thickness and thus an oval disc like gemma is formed.
(ix) Thallus of Anthoceros
External features : The plant body is small, prostrate, dorsiventral spongy thallus of dark-green colour. There is no conspicuous midrib although the middle part of thallus is many layered. The dorsal surface of the thallus may be smooth or rough with ridges or even with some lamellar growth. The lower ventral surface bears numerous smooth-walled rhizoids. Scales or mucilage hairs are absent. Several large dark bluish green patches of Nostoc colonies can be seen on the ventral surface of thallus. In A. hallii the thallus is long and pinnately compound.
Internal features : The thallus shows little or no cellular differentiation. All the cells are alike except the upper epidermis with comparatively smaller cells. The thallus is composed of uniform, thin-walled, parenchymatous cells. Each cell of the thallus shows a single oval chloroplast with a central pyrenoid. The air-chambers or pores are absent but the lower surface has intercellular cavities filled with mucilage.

The mucilage cavities always contain Nostoc colonies which do the function of nitrogen-fixation. The mucilage cavities open on the ventral surface through slime pores. The thickness of the thallus may vary as 6-8 cells in A. laevis or 30-40 cell in A. crispus.
The growth of the thallus takes place by the activity of a single apical cell with two cutting faces. But A. himalayensis grows by a group of apical cells. The apical cell may split into two giving rise to the dichotomous lobes.
(x) Rhizome of Ophioglossum
A transverse section of the rhizome reveals an irregular outline due to the attachment of adventitious roots and leaf bases. The cortex remains bounded by a single-layered epidermis. The cortex consists of large thinwalled parenchymatous cells having abundant intercellular spaces among them. Maheshwari and Singh reported the formation of a periderm layer in old rhizomes; but there is no functional cambium.

The lowermost portion of the rhizome may have an endodermis delimiting stele and cortex (Bower). In upper portion of the rhizome there is no endodermis. Here the innermost cells of the cortex and the outermost cells of the stele are so similar in appearance that it becomes difficult to differentiate where one region leaves off and the other begins. Lang has reported an internal and external endodermis in young plants of a Ceylonese species of Ophioglossum.
Generally the rhizome possesses an ectophloic siphonostele throughout its entire length. This siphonostele may have but one leaf gap when cut at any particular level. More often two or more leaf gaps may overlap forming a dictyostelic condition. Most of the species have a single leaf trace departing at each gap and traversing the cortex without branching. A few species, e.g. O. palmatum and O. pendulum have two traces departing at each gap. There is a division of each trace into several strands before it runs into the leaf base.
The xylem of each meristele is endarch in arrangement. The protoxylem remains situated on the inner face of the xylem mass. The reticulate tracheids of irregular shape form the metaxylem. Boodle (1899) has reported a small amount of secondary thickening in the stem of O. vulgatum. The protoxylem is composed of spiral and scalariform elements. In O. palmatum, the protoxylem is ill defined. The relation of the xylem to the phloem is collateral, that is the phloem lies external to the xylem. Commonly the phloem consists of a layer of four or five cells in thickness. However, in O. pendulum the phloem consists of a single layer of cells separated from the xylem by a layer of parenchyma three to five cells in thickness. The pith internal to the xylem is composed of thin walled large parenchymatous cells. According to Petry, sometimes (e.g., in O. pendulum) the tracheids occur in the pith region.
(xi) Heterospory
The sporangia show greater specialization than those of ferns. They are differentiated into micro and megasporangia. The microsporangia contain microspores whereas megasporangia contain megaspores in them. Each microsporangium contains a large number of microspores whereas each megasporangium contains usually only four megaspores. The condition of the production of two kinds of dissimilar spores, differing in shape and size in the same species, is termed heterospory. When one kind of spores are present, e.g., Equisetum and ferns, the condition is said to be homospory. According to modern pteridologists the heterospory is found in seven genera-Selaginella, Isoetes, Marsilea, Salvinia, Azolla, Regnellidium and Pilularia. Here one thing may be noticed that the microspores develop male gametophytes whereas the megaspores germinate into the female gametophytes which may also be called as microgametophytes and megagametophytes. In the case of homospory the sex may be differentiated only at the gametophytic stage, but here the sex may be differentiated even at the sporophytic stage by the presence of small and large sized microspores and megaspores which produce antherozoids and eggs respectively.
The differentiation of microspores and megaspores and their dependence upon the sporophyte has certain advantages. The gametophytes of the ferns are, however dependent for their nutrition upon soil and environmental conditions, whereas, in the case of Selaginella, as far as the nutrition of gametophytes is concerned they derive it from the sporophyte, and therefore they are more independent to the external conditions than those of ferns.
The condition of heterospory in Selaginella constitutes one of the most important phases in the evolution of the plants. In certain species of Selaginella only one megaspore develops within the megasporangium. The highest evolved species i.e., S. apoda has advanced almost upto the rank of a seed plant. Here, the megaspore remains within the sporangium; it itself fertilizes and this way for the first time the gametophyte of Selaginella shows complete dependence of the gametophyte upon sporophyte as in angiosperms.
Biological Importance of Heterospory: The phenomenon of heterospory is of great biological advantage because of the fact that a large megaspore which contains female gametophyte derives its food from the sporophyte, and is independent of the external conditions as might interfere with the growth of a free living gametophyte. It thus forms better starting point for the new embryo, than an independent green prothallus which has to manufacture its own food.
In brief, the origin of seed habit is associated with the following important prerequisites :
1. The production of two kinds of spores (i.e., heterospory).
2. The retention and germination of the fertilization of the egg and embryo formation, which still within germinating megaspore.
3. Development of only one megaspore per megasporangium. In Selaginella, there is remarkable approach to the seed habit, on account of the following important features :
(i) Selaginella shows heterospory.
(ii) The megaspore usually germinates within the megasporangium and is not shed for a time which may vary from species to species.
(iii) There is reduction to one megaspore in some species, e.g., S. rupestris and S. monospora and a confirmed tendency to reduction in others.
It becomes quite evident that Selaginella has considerably advanced towards the seed habit in a few species, but its approach to the true seed is not complete due to the following features :
(i) The megasporangium lacks an integument or covering.
(ii) The permanent retention of the megaspore within the megasporangium has not become established.
(iii) After the development of the embryo, the resting period is not there.
(xii) Protostele
Protostele, according to Jeffry (1903) and most of other present day workers, is the most primitive and simplest type of stele. It consists of solid mass of xylem, completely surrounded by phloem, pericycle and endodermis in regular order (Fig.). Protostele is permanently remained in the adult stems of many living pteridophytes e.g. Lygodium, Lycopodium, Hymenophyllum, Selaginella etc.
The protostele is the fundamental types of stele for the vascular plants and all others steles have been derived from it during the course of evolution.
The protostele may exist in any of the following form :
1. Haplostele: Haplostele is that in which the central xylem has a smooth outslide and surrounded by concentric layers of phloem e.g., Lygodium, Selaginella, kraussiana and sps. of Lycopodium. This Haplostele is regarded as the most primitive of all the protostele.
2. Actinostele: Actinostele is that in which the central xylem cone has radiating ribs and assumes a star shaped appearance e.g. Lycopodium, C. serratum, Psilotum.
3. Plectostele: Plectostele is that in which the Xylem is divided into a number of separate plates, which lie parallel to or radiate to one another e.g., Lycopodium volubile.
4. Mixed protostele : Mixed protostele is that in which the Xylem and phloem are uniformly distributed. Xylem elements remain embedded in phloem tissue, e.g., Lycopodium cernuum. The stele is without pith.

Fig. A, B, C, D-Showing various types of protostele (Diagrammatic). A-Haplostele, B-Actinostele, C-Plectostele, D-Mixed protostele.
(xiii) Types of Siphonostele or Medullated Protostele
Siphonostele is that stele in which the pith is present in the centre of hollow vascular cylinder. So siphonostele is actually a modified protostele with pith and therefore botanists believe that siphonostele has arising from protostele either by the intrastelar origin of pith or by the invasion of cortical tissue (extrasteler origin of pith). The first view, which has been proposed by Bordle, has been accepted by most of the workers.
Siphonosteles may of the following three types:
1. Ectophloic siphonostele: This types of stele has central pith, which is surrounded by concentric rings of xylem and phloem.
2. Amphiphloic siphonostele : In this type of stele the centre i is occupied by pith. In this case the xylem is surrounded on both external and inner sides by the rings of phloem, pericycle and endodermis, e.g. Marsilea.
3. Eustelic siphonostele: In this type of stele, the xylem and phloem cylinders breaks into a large number of collateral vascular bundles, which are arranged in a ring e.g. Equisetum.

Fig. A, B, C- Various types of Siphonosteles.
A-Ectophloic, B-Amphiphloic, C-Eustelic (Diagrammatic)
Solenostele: Solenostele is that stele in which the stele is perforated by single leaf. It may be ectophloic or amphiphloic or solenostele in the same way as in siphonostele e.g. ferns (Fig.).
Dictyostele: Dictyostele is that stele in which there occur more than one leaf gap. The portion of vascular tissue which lies in between two gaps is termed as Meristeles. Each Meristele is protostelic in nature, e.g., Ferns.
Polycyclic stele: Polycyclic stele is that stele in which the meristeles are distributed in more than one rings and are of different sizes. There occur normally two rings in Pteridium aquilinum (fern).
(xiv) Stem anatomy of Lycopodium
The stem displays its interesting anatomical features specifically with regard to stelar organization. The outline in T. S. is not perfectly cylindrical owing to the presence of closely-placed leaf-bases.
Epidermis: It is uniseriate parenchymatous bearing a cuticle. The outer wall of this layer is cutinized,
Cortex: The broad multilayered cortex lying in between epidermis and endodermis is either homogeneous or heterogeneous. In L. selago, the homogeneous parenchymatous cortex has intercellular spaces; however its mature stem is purely sclerenchymatous throughout.
The heterogeneous cortex is divisible in three distinct zones i.e., in L. clavatum, both the outer and inner zones are multilayered sclerenchymatous, while the middle zone being multilayered parenchymatous bearing the bundles of leaf traces; however just the reverse condition is met with in L. cernuum having the mid zone sclerenchymatous while the outer and inner parenchymatous.
Endodermis and Pericycle: Both individually are uniseriate. The endodermal cells with casparian strips become thickened in mature stem.
Vascular cylinder: It is the simplest protostele. Based on the position and arrangement of vascular tissues, that are accounted in different species, altogether there are four kinds of protostele.
In those cases where the stems are relatively bulky, the central part of mixed protostele has sufficient parenchymatous cells.
In all the aforesaid instances the protoxylem elements are exarch.
The xylem is mainly composed of tracheids. The protoxylem has got annular or spiral thickenings, while the metaxylem in its tracheidal walls carries either circular pits or scalariform thickenings. The phloem, on the other hand, comprises mainly the sieve tubès and phloem parenchyma.
The most primitive protostele is a fundamental vascular structure for Pteridophytes, and all other types of steles have been derived from it during the course of specialization.
(xv) Sporangia of Rhynia
The sporangia were cylindrical and borne singly on the tips of some aerial branches. They were large, oval or cylindrical structures with pointed ends. The sporangia had walls several cell layers thick in which the cells of the outermost layer were thick walled and had a heavy cuticle. The sporangium was without any specialised mechanism of dehiscence. The inner cells of the jacket probably functioned as tapetum. The sporangia contained numerous spores with cutinised walls. The spores were apparently all alike and were arranged in tetrads. The presence of tetrads in some specimens suggests that meiosis occurred in Rhynia and that the plant body, therefore, belonged to the sporophytic generation.
(xvi) L. S. Capsule of Sphagnum
It is spherical and dark brown in colour. It contains central columella which is over arched by a dome shaped spore sac containing haploid spores. The wall of the capsule is 4-6 layers thick. The outer layer of the capsule wall is called epidermis. The cells of the epidermis are compactly arranged and contain chloroplasts.
It has many non-functional and rudimentary stomata. In young sporogonium, a circular, convex disc is present at the top of the capsule. It is called operculum. It is separated from the rest of the capsule region by a circular (ring like) groove of thin walled cells called annulus.
On sunny days the columella of mature capsule dries up, breaks down and forms a large air cavity below the air sac. Air enters in this cavity through the rudimentary stomata.
The wall of the capsule dries up under the influence of the sun and spherical form of the capsule gradually becomes cylindrical. The imprisoned air in the capsule is compressed and held under considerable pressure due to change in the shape of the capsule.
The operculum also dries and shrinks. Due to this a little difference in the tension is set up. It puts a strain on the thin celled annulus cell which finally rupture under the mounting pressure of imprisoned air. The operculum is blown off with explosive force.
The pressure of imprisoned air also ruptures the spore sac and the cloud of yellow or orange coloured spores is blown to a height of several centimeters in the atmosphere. method of spore discharge is, therefore, known as the air gun or explosive mechanism.

(xvii) Economic importance of Sphagnum
Sphagnum is known as a ‘peat’ or ‘bog moss’ which is of commercial value in horticulture because of its high water retaining capacity. It alongwith other plant remains increases the acidity of soil even when it is dead and pulverized. It inhabits water and forms extensive surface mats or ‘quacking bogs’.
Acidic nature of bogs prevents the growth of micro-organisms including Bacteria, so the constant accumulation of Sphagnum (due to death and decay of stem axes) results in forming the deep deposits called peat. The lower strata of such deposits may be of considerable age, for instance some bogs in U.S.A. are 16,000 years old. Practically the peat is used in improving the texture and water holding capacity of soils, and in providing nutrients to cultivated plants. In some parts of world where prevails the deficiency of coal, the peat is compressed, dried and burned. The flavour of ‘Scotch whiskeys’, in part, is due to peats smoke.
It acts as an insulator against heat and cold during shipment. By virtue of its antiseptic property, it was used in surgical dressing (in place of cotton) during World War I and Russio-Japanese war.
It is immensely used in garden beds for proper germination of seeds and in green houses for raising the plants from cutting.
Owing to its spongy texture coupled with high absorptive power, it is used for packing the bulbs, seedlings, cuttings, and for a lot of other materials where moisture is imperative. Glassware equipments are packed by it while they are exported.
(xviii) Cone of Calamites
The cone of calamites has a central axis bearing alternating whorls of peltate sporangiophores and sterile appendages called bracts. There are a number of different forms of cones in calamites. These include
(i) Palaeostachya: The sporangiophore whorls alternate with sterile bracts whorls. The sporangiophore whorls arise from the axils of bracts and have independent vascular supply-

(ii) Calamostachys : The sporangiophore whorls and bracts whorls are similarly alternating with each other but both whorls arise directly from the stem axis at right angles to it so that they look quite independent. The sterile bracts are found laterally at the base. There is a variation in the number of sporangiophores and bracts. Usually four sporangia are borne at the peltate disc of sporangiophore. The sporangia contain a large number of spores. The spores are normally similar in size presenting a homo-sporous condition but in some species (C. casheana) the heterosporous condition is seen.
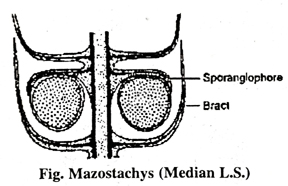
(iii) Mazostachys Here sporangiophores are borne in a whorl just below the vertical of bracts. The ratio of bracts to sporangiophores is 2 : 1, where a whorl of 12 bracts subtended by a whorl of 6 sporangiophores. The sporangiophores bear two pendant sporangia. The sporangiophore trace directly from the node before bending outward into the sporangiophore.
(iv) Cingularia: It resembles Mazostachys. Here both the whorls of bracts and sporangiophores are fused and develop horizontally. The sporangiophores are flat and bifurcate at their tips bearing four pendant sporangia.
(v) Calamocarpon : It is the most highly evolved cone among calamites. In general organisation, calamocarpon resembles calamostachys. The bracts sporangiophores ratio is 1 : 1. There are variable numbers of bracts per to whorl.
(xix) Protocorm
Protocorm is a tuberous mass of cells that is the embryonic form of orchids and club mosses.
Cernuum and other forms with superficial green prothalli is attached to the prothallus by a small foot, and develops at first as a tuberous body known as protocorm bearing rhizoids. This forms a number of simple leaves, and upon it the apex of the shoot arises later.
When new individuals of species which possess a protocorm arise vegetatively from the leaves or roots of young plants, the protocorm appears in the young sporophyte. The plant is reproduced by tubers, which resemble the protocorm, bearing first a number of protophylls and later the upright shoot with its single terminal strobilus.
(xx) Sporangia of Selaginella
Each sporophyll bears a single stalked sporangium situated in the axil of it, on the adaxial side between ligule and its base. There are two kinds of sporangia. The sporangia bearing megaspores are called megasporangia and the sporangia with microspores are microsporangia. In the same way the sporophylls bearing megasporangia are megasporophylls and bearing micro sporangia are microsporophylls.
The development of mega and microsporangia is quite common upto the formation of spore mother cells. There are two schools of thought about the nature of their development. According to one school of thought the sporangium develops from a single initial cell. According to other school, the sporangium arises from a transverse row of initials. In majority of the species the sporangia arise from the cells of the axis and said to be cauline whereas Bower holds the view that the sporangium arises from the base of the sporophyll and foliar in origin. Any how, the mature sporangium is a distinctly axillary structure at the base of the sporophyll.
The sporangia which are to develop into microsporangia contain all the sporogenous cells. The sporogenous cells of the last generation are the sporocytes. During this process 10 to 20 percent of the sporogenous cells are disintegrated. This way the sporocytes become loosened within the sporangial chamber. The functional sporocytes become rounded and separated from each other. Each sporocyte (spore mother cell) divides meiotically giving rise to a tetrad of four spores. This way numerous haploid microspores are developed within the sporangium.
The sporangia which are to develop into megasporangia, contain many sporocytes in them in the beginning. Later on all sporocytes but one is disintegrated. The functional megaspore mother cell undergoes the usual reduction division and a tetrad of megaspores is developed.
Follow on Facebook page – Click Here
Google News join in – Click Here
Read More Asia News – Click Here
Read More Sports News – Click Here
Read More Crypto News – Click Here